|
УДК 594.3 Л.М. Хлус,
кандидат біологічних наук, доцент
(Чернівецький національний університет);
К.М. Хлус,
кандидат біологічних наук, зав. лабораторією
(Науково-дослідний інститут медико-екологічних проблем, м. Чернівці) Конхологічна мінливість молюска Eobania vermiculata Müll. в онтогенезі Вивчено зміни конхологічних параметрів наземного молюска Eobania vermiculata Müll. в онтогенезі. Виявлено відмінності ступеня наростання базових метричних показників при зменшенні кулястості черепашки. Припускається адаптивний характер явища у зв’язку з аридністю клімату. Об’єктом даного дослідження було обрано онтогенетичні зміни конхологічних метричних параметрів наземного молюска еобанії Eobania vermiculata Müll. (Gastropoda, Pulmonata, Geophila, Helicidae). Вид широко розповсюджений у країнах Середземномор’я; окрім того, він був інтродукований у Північну Америку [1]. Нещодавно цей слимак був знайдений в Середній Азії. Вважається, що він завезений туди колекціонерами-любителями [2]. Припускають, що під час російсько-турецьких війн (середина ХІХ ст.) E.vermiculata була завезена у Крим; вперше була виявлена в районі міста Севастополь [3], де успішно акліматизувалася і почала розселюватися по території півострова [4]. Існує думка, згідно якої вид був завезений, можливо, ще у період колонізації греками або ж генуезцями як їстівна тварина, призначена для розведення [5]. Нині вид виявлений повсюдно у прибережній зоні, в центральному Криму, в зоні передгір’я та на Керченському півострові [6]. Як це часто буває на межах ареалу (в даному випадку північних), у Криму еобанія виражено тяжіє до антропогенних ландшафтів, стаючи типовим синантропним видом. Молюски зустрічаються у садах, прибережних чагарниках та рідкому деревостані, вздовж парканів на вузькій смузі узбережжя у великих куротних населених пунктах південно-східного Криму (у Феодосії, Судаку тощо), у зв’язку з чим можуть бути використані для біомоніторингу довкілля в зонах високого рекреаційного навантаження [7].
В той же час стан вивчення біології цього молюска можна вважати незадовільним. Серед нечисельних робіт можна вказати на матеріали досліджень анатомії статевої системи [8], характеру добової локомоторної активності E.vermiculata в аспекті аналізу біоенергетики виду [9], репродукційних властивостей та особливостей ювенільного росту цих молюсків [6].
Черепашка E.vermiculata має характерну для геліцид форму: від низько кубареподібної до притиснуто-конічної, з тупим конічним завитком, висота якого більша від висоти устя [1 :10]. Вона товстостінна і міцна, сірувато-біла або жовта, одноколірна або з 4-5 спіральними коричневими смужками, які інколи зливаються одна з одною, а інколи розпадаються на ряди плям. Верхня смуга – найбільш широка – лежить безпосередньо під швом, друга і третя смуги часто зливаються одна з одною. Поверхня черепашки неясно поперечно покреслена і покрита чисельними білими короткими поздовжніми зморшками, які часто переплітаються одна з одною і утворюють неправильну сітку. Обертів 5, слабо опуклих, плавно і швидко наростаючих, ембріональні оберти (1¼) гладенькі, останній оберт сильно здутий, округлий, біля устя в 1½ - 2 рази ширше передостаннього і різко опущений. Устя короткоовальне, дуже косе, всередині біле, краї його гострі, сильно вивернуті і потовщені білою губою; колумелярний край широкий, блискучий, на всій відстані щільно злитий з нижньою стороною черепашки. Пупка немає. Висота черепашки 17-22 см; ширина 27-32 мм.
Відомості щодо особливостей ростових процесів в онтогенезі еобаній у періодичній та монографічній літературі поодинокі. Так, у недавньому дослідженні [6] розглянуті особливості репродукції цих тварин. Показано, що середні розміри кладки для виборок з п’яти популяцій в цілому подібні і лише незначно відрізнялися від відповідних показників для тварин з Греції. В той же час, мінливість цієї ознаки досить висока. Виявлена також мінливість розмірів яєць як між особинами з різних популяцій, так і у межах однієї кладки для особин з однієї популяції; вважається, що вона відображає генетичну неоднорідність особин. Відомо, що швидкість статевого дозрівання гелікоідей тісно пов’язана з тривалістю їх життя. Молюски цієї групи можуть мати одно-, двох-, трирічний та багаторічний цикли [11; 12]. Даних щодо тривалості життя E.vermiculata в літературі не знайдено, проте взагалі у молюсків вона тісно пов’язана з розміром: великі молюски, схожі за розмірами на еобанію, живуть по багато років, у зв’язку з чим розмноження в них починається лише на 3-4-му році життя [1]. Так, равлики весняної генерації Bradybaena fruticum Müll., які досліджувалась на східній межі свого ареалу (р.Об), стають статевозрілими на четвертому році життя, протягом п’ятого вони є репродуктивною групою і навесні шостого року гинуть [13]. Виноградні слимаки [14] в умовах Литви починають розмножуватися на 4-му році життя.
Виходячи з викладеного вище, можна було припустити, що мінливість ознак черепашки у еобаній різних вікових груп може відрізнятися, відображаючи особливості ростових процесів на окремих етапах онтогенезу. Оскільки відомо, що швидкість росту черепашки наземних молюсків, як і її мінливість, помітно впливає на характер динаміки чисельності та розмірно-вікового складу популяції [15; 16], вивчення цих процесів може логічно доповнити загальну картину внутрішньопопуляційної та міжпопуляційної мінливості еобаній, раніше продемонстровану нами [7]. Для вивчення вікових особливостей ростових процесів E.vermiculata вибірку тварин з м. Феодосії (район Карусельної гірки) розділили на 2 вікові категорії – статевозрілих (зі сформованою губою устя) та нестатевозрілих. Для кожної з них був проведений весь блок морфометричних досліджень: визначали висоту черепашки (ВЧ) і вустя (ВВ), великий (ВД) і малий (МД) діаметри, ширину вустя (ШВ) і кількість обертів (КО), розраховували морфометричні індекси як співвідношення вказаних вище метричних показників; основні результати цієї роботи наведено раніше [7]. Зокрема, показано, що у статевозрілих тварин зростають всі лінійні параметри, у тому числі і кількість обертів, а з морфологічних індексів змінюються лише ті, що характеризують стан радіальних параметрів (ВД та МД) та їх співвідношення з іншими, а саме ВЧ/ВД, ВЧ/МД, ВВ/ВД, ВВ/МД, ШВ/ВД (рисунок). 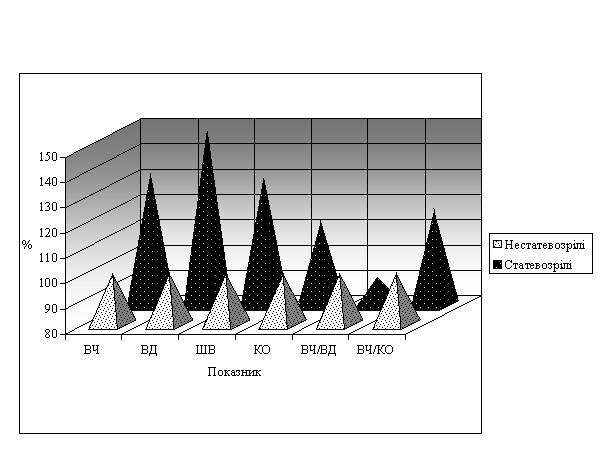
Рис.1. Онтогенетичні зміни морфологічних параметрів черепашок Eobania vermiculata Müll. Ці факти вказують на кінцеву детермінованість ростових процесів переважно за рахунок останнього оберту. Враховуючи, що, як в екофізіологічних, так і в популяційних дослідженнях, більшу цінність та інформативність мають не самі показники, а швидкість та/або інтенсивність їх змін [17], ми обчислили прирости лінійних параметрів черепашки за різницею відповідних значень морфометричних показників в групах статевозрілих і нестатевозрілих тварин (dР) відносно приростів кількості обертів (dKO), позначивши це відношення як ступінь наростання параметру, і отримали наступні результати: dВЧ –––– =10,17; dКО | dВД –––– =18,67; dКО | dМД –––– =14,75; dКО | dВВ –––– =6,08; dКО | dШВ –––– =7,40; dКО |
Приріст габітуальних ознак, особливо ВД і МД, виявився значно більшим, ніж відповідний показник для параметрів устя, що природним чином відображає залежність ступеня збільшення остаточних габітуальних показників від збільшення КО на останньому році життя. Для усунення можливого впливу більших абсолютних значень лінійних розмірів у порівнянні з розмірами устя, наведений показник модифікували, використавши замість приросту лінійних параметрів (dP) їх відносний приріст (dP/P), виразили його у відсотках і також віднесли до змін кількості обертів (dКО): dВЧ –––––––– · 100 = 65,51; ВЧ · dКО | dВД ––––––– · 100 =101,85; ВД · dКО |
dМД –––––––– · 100 =90,49; МД · dКО | dВВ –––––––– · 100 =66,74; ВВ · dКО | dШВ –––––––– · 100 =62,68; ШВ · dКО |
З наведених результатів видно, що осьовий габітуальний показник ВЧ значно менше реагує на наростання останнього оберту, ніж радіальні параметри. Отже, збільшення розмірів черепашки E.vermiculata в онтогенезі веде до зміни її форми з більш кулястої до все більш присадкуватої. Беручи до уваги особливості впливу кліматичної температури на загальну форму черепашки (еколого-еволюційне правило Хеммінгсена щодо розмірів черепашок наземних молюсків) [18], можна припустити, що у нестатевозрілих молюсків менша відносно об’єму тіла його поверхня запобігає надмірному нагріванню і втраті організмом води в умовах сухого спекотного клімату м. Феодосії (середньорічна кількість опадів 376 мм, середня температура липня 23,8оС, а максимальна перевищує 40 оС) [19]. Дорослі ж тварини, завдяки великим розмірам та товстішій черепашці, менше потерпають від проявів аридності клімату, що і відбивається на формі черепашки. З іншого боку, присадкувата форма вигідніша з точки зору використання тимчасових схованок від денної спеки, а як такі еобанії використовують прикореневі частини рослин, щілини у кам’яних кладках та бетонних мурах, заповзають під каміння тощо. Цікаво також, що форма устя в онтогенезі залишається незмінною.
Отже, значне збільшення розмірів черепашки E.vermiculata Müll. в онтогенезі відбувається при диференційних ступенях наростання окремих конхологічних параметрів. Габітуальні особливості кожної вікової категорії мають адаптивний характер у зв’язку з високим ступенем аридності досліджуваного біотопу. СПИСОК ВИКОРИСТАНОЇ ЛІТЕРАТУРИ 1. Шилейко А.А. Наземные моллюски надсемейства Helicoidea // Фауна СССР. Моллюски. – Л.: Наука, 1978. – Т.3, вып.6. – 384 с.
2. Иззатуллаев З.И. О новом завезенном в Среднюю Азию наземном моллюске Eobania vermiculata (Pulmonata, Helicidae) // Зоол. журн. – 1996. – 75. – С. 778-780.
3. Rosen O. Helix vermiculata Müll. in der Krim // Nachrbl. Malak. Ges. – 1911. – B. XLIII. – S. 32.
4. Пузанов И.И. Материалы к познанию наземных моллюсков Крыма, ч. III. Состав, распределение и генезис Крымской малакофауны // Бюлл. Московск. общ. испыт. прир., отд. биолог. - 1927. – Т. XXXIV. – С. 221-282.
5. Лихарев И.И. Некоторые факторы, определяющие распространение наземных моллюсков // Моллюски. Вопросы теоретической и прикладной малакологии. – М. – Л.: Наука, 1965. – С. 48-51.
6. Крамаренко С.С., Попов В.Н. Особенности репродукции и роста наземного моллюска Eobania vermiculata (Müller, 1774) (Gastropoda; Pulmonata; Helicidae) в лабораторных условиях // Экология. – 1999. – N 4. – С. 229-302.
7. Хлус Л.М., Хлус К.М. Морфологічні параметри Eobania vermiculata Müll. (Gastropoda, Helicidae) як індикатор рекреаційного навантаження на екосистеми південного сходу Кримського півострова // National natural parks: Problems of creation and development. – Proceedings of the international scientific-practical conference, decated to the 20th-anniversary of the Carpathian national natural park. – September 14-17, 2000. – Yremche, Ukraine, 2000. – P. 338-341.
8. Арутюнова Л.Д. Заметки о некоторых наземных моллюсках южнобережья Крыма // Биол. журн. Армении. – 1975. – 28. – С.104-109.
9. Зейферт В.Д. Суточная активность и характер перемещения наземного моллюска Eobania vermiculata (Müller) // Экологическая энергетика животных. – Свердловск, 1988. – С. 125-134.
10. Лихарев И.М., Раммельмейер Е.С. Наземные моллюски фауны СССР. – М. – Л.: Изд. Московского университета, 1952. – 512 с.
11. Deblock R., Hoestland H. Donnes biologiques le Gastropoda litorae Theba pisana Müller aux limites septentrionales de "son axtention" // C. r. Acad. sci. – 1967. – V. 265. – N 12. – P. 893-896.
12. Chatfield J.E. The life history of the heliciol snail Monacha cantiana (Montagu), with reference also to M.carthucha cantiana (Müller) // Proc. Malacol.Soc.London. – 1968. – V. 38. – N 3. – P. 233-245.
13. Зейферт Д.В. Некоторые аспекты биоэнергетики наземных моллюсков // Энергетика роста и развития животных. – Свердловск: УНЦ АН СССР, 1985. – С. 76-88.
14. Андрейкович Е.В. О размножении Helix pomatia L. в Литовской ССР // Моллюски и их роль в экосистемах. – Л, 1968. – С. 59-61.
15. Oosterhoff L.M. Variation in growth rate as an ecological factor in the landsnail Capaea nemorales (L.) // Neth. J. Zool. – 1977. – V. 27. – P. 1-132.
16. Heller J., Jttiel H. Natural history and population dynamics of the land snail Helix texta in Israel (Pulmonata; Helicidae) // J. Mollusc. Stud. – 1990. – V. 56. – P. 189-204.
17. Шилов А.И. Экология. – М.: Высш. шк., 1997. – 512 с.
18. Терентьев В.П. Влияние климатической температуры на размеры раковины наземных моллюсков // Зоол. журнал. – 1970. – Т. XLIX. – Вып. 1. – С. 5-10.
19. Подгородецкий П.Д. Крым: Природа: Справочное издание. – Симферополь: Таврия, 1988. – 192 с. Матеріал надійшов до редакції 27.06.01. Хлус Л.Н., Хлус К.Н. Конхологическая изменчивость моллюска Eobania vermiculata Müll. в онтогенезе.
Изучены изменения конхологических параметров наземного моллюска Eobania vermiculata Müll. в онтогенезе. Выявлена разница степени нарастания базовых метрических показателей при уменьшении шаровидности раковины. Предполагается адаптивность явления в связи с аридностью климата. Khlus L.M., Khlus K.M. Conchological variation of mollusk Eobania vermiculata Müll. for ontogenesis.
The difference of the degree of increase of basic metric indices at decrease of shell’s globularity is determined. The phenomenon adaptivity due to the arid character of the climate is suggested. Всі опубліковані на сайті матеріали належать їх авторам. Матеріали розміщено виключно для ознайомлення. Копіювання та використання інформації суворо заборонено.
|







